答(da)案可能是NO.
我們是多(duo)民族(zu)融(rong)合而成的么(me)?
答(da)案可能還是NO.
我們(men)是誰,我們(men)從何(he)而來?
2003年4月14日,美國科學家在(zai)(zai)華盛頓莊(zhuang)嚴(yan)宣布:美、英、日、法、德和中國,6個國家聯合,經過13年的(de)努力,共(gong)同繪制完(wan)成(cheng)了《人類基(ji)因序(xu)列圖》。由IBM贊助,美國《國家地(di)理(li)》雜志舉行的(de)研究(jiu)(jiu)“人類遷徙遺傳地(�������di)理(li)圖譜計劃”2005年4月開始在(zai)(zai)世界各地(di)進(jin)行,復旦大學生命(ming)科學院承(cheng)擔了遠東地(di)區及其東南亞(ya)地(di)區的(de)DNA的(de)取樣和研究(jiu)(jiu)。
隨著(zhu)研(yan)究的深入,答案漸漸浮出水(shui)面,一張完整的人類(lei)遷移路(lu)線圖(tu)開(kai)始(shi)顯現。我們(men)整理了(le)一些大(da)家(jia)比較關�����注的內容。
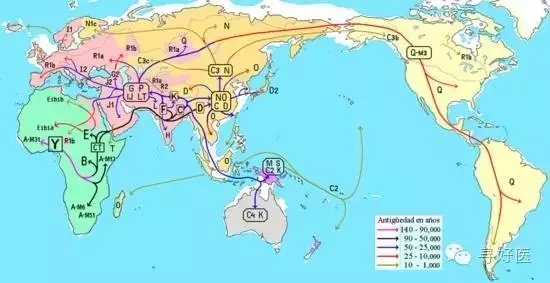
全(qua������������n)世界人口基因來(lai)自(zi)36個(ge)女(nv)人
源(yuan)于(yu)“線粒(li)體夏娃”
令人(ren)(ren)(ren)(ren)意(yi)想(xiang)不(bu)到(dao)(dao)的(de)是我們60多億人(ren)(ren)(ren)(ren)口可(ke)能源(yuan)自(zi)一(yi)個(ge)(ge)(ge)母(mu�������)(mu)(mu)親。英國牛津大(da)學人(ren)(ren)(ren)(ren)類(lei)遺傳學家(jia)經十幾年(nian)的(de)DNA研(yan)究發現,全世界(jie)的(de)人(ren)(ren)(ren)(ren)口分(fen)別(bie)繁(fan)衍自(zi)36個(ge)(ge)(ge)不(bu)同的(de)、被稱做“宗族(zu)母(mu)(mu)(mu)親”的(de)原始女人(ren)(ren)(ren)(ren),其中非(fei)洲有13個(ge)(ge)(ge)“宗族(zu)母(mu)(mu)(mu)親”,印度、澳大(da)利亞、中亞有7個(ge)(ge)(ge)。而所有這些(xie)“宗族(zu)母(mu)(mu)(mu)親”又都是15萬(wan)年(nian)前到(dao)(dao)20萬(wan)年(nian)前非(fei)洲大(da)陸上一(yi)個(ge)(ge)(ge)科(ke)學家(jia)命名(ming)為“線粒體夏娃”(Mitochondrial Eve)的(de)女人(ren)(ren)(ren)(ren)的(de)后代。盡管“夏娃”不(bu)是當時(shi)惟一(yi)活(huo)著(zhu)的(de)女性,然(ran)而她卻是惟一(yi)一(yi)個(ge)(ge)(ge)將血脈延續繁(fan)衍到(dao)(dao)今天的(de)原始女人(ren)(ren)(ren)(ren)。
據(ju)英(ying)(ying)國(guo)《每日快報》報道(dao),該理論是(shi)由英(ying)(ying)國(guo)牛(niu)(niu)津大學人類遺(yi)傳學教(jiao)授布賴(lai)恩·西基斯(Bryan Sykes)提(ti)出的(de),他是(shi)世(sh����i)界上(shang)第一個(ge)發明從年(nian)代(dai)久遠的(de)古代(dai)骨骼中提(ti)取出DNA方法的(de)遺(yi)傳學者,并建立了“牛(niu)(niu)津祖先”(Oxford Ancestors)項目。
西基(ji)斯教授研究發現,現代歐洲人(ren)(ren)其(qi)實(shi)(shi)大多數都(dou)是(shi)遠親:97%的(de)(de)現代歐洲人(ren)(ren),其(qi)實(shi)(shi)都(dou)起源于10000年前(qian)到(dao)45000年前(qian)冰河時代的(de)(de)7個(ge)不同(tong)女(nv)人(ren)(ren),這7個(ge)“宗族母(mu)(mu)親”被(bei)他稱(cheng)做是(shi)“夏(xia)娃的(de)(de)7個(ge)女(nv)兒(er)”,7名原始女(������nv)人(ren)(ren)通(tong)過“線(xian)粒(li)體(t�����i)DNA”和現代歐洲人(ren)(ren)聯系(xi)(xi)到(dao)了一起。線(xian)粒(li)體(ti)DNA是(shi)只(zhi)通(tong)過母(mu)(mu)系(xi)(xi)一脈傳遞的(de)(de)遺(yi)傳基(ji)因(yin),男性也能從母(mu)(mu)親那(nei)(nei)(nei)里繼承線(xian)粒(li)體(ti)DNA,但卻無法將(jiang)它(ta)遺(yi)傳給(gei)自己的(de)(de)后代。也就是(shi)說,如果一個(ge)女(nv)性生(sheng)下的(de)(de)全都(dou)是(shi)兒(er)子,那(nei)(nei)(nei)么她的(de)(de)線(xian)粒(li)體(ti)DNA遺(yi)傳鏈將(jiang)因(yin)此終止。所以,如果一個(ge)女(nv)人(ren)(ren)沒生(sheng)出女(nv)兒(er),那(nei)(nei)(nei)她就真的(de)(de)斷子絕孫了。
西(xi)基斯稱(cheng),“很(hen)顯(xian)然(ran),在漫長(chang)的(de)冰河時代(dai),歐(ou)洲并非只生(sheng)存著這(zhe)(zhe)7個(ge)(ge)女(nv)人(ren),同(tong)一(yi)時代(dai)生(sheng)活著大(da)量(liang)的(de)原(yuan)始(shi)(shi)女(nv)性(xing)。然(ran)而她們要(yao)么沒(mei)有(you)長(chang)到(dao)成年(nian)(nian),要(yao)么沒(mei)有(you)生(sheng)下孩(hai)子,要(yao)么生(sheng)下的(de)全是男孩(hai),但這(zhe)(zhe)7個(ge)(ge)原(yuan)始(shi)�������(shi)女(nv)人(ren)不(bu)僅活了(le)足夠(gou)長(chang)時間,并且(qie)(qie)每人(ren)至(zhi)少生(sheng)了(le)兩名女(nv)兒,從而開始(shi)(shi)了(le)線(xian)粒體DNA遺傳鏈,并且(qie)(qie)一(yi)直延續到(dao)了(le)今天。 據西(xi)基斯教(jiao)授稱(cheng),線(xian)粒體DNA一(yi)般很(hen)難發生(sheng)改變,平均要(yao)過2萬年(nian)(nian)線(xian)粒體DNA才會發生(sheng)微小(xiao)的(de)變異(yi)。他(ta)對記者道:“研究發現,大(da)多數(shu)現代(dai)歐(ou)洲人(ren)的(de)線(xian)粒體DNA可(ke)分為7種類(lei)型,就仿佛血(xue)型一(yi)樣。我們相信,每個(ge)(ge)線(xian)粒體DNA相同(tong)的(de)人(ren)都是數(shu)萬年(nian)(nian)前同(tong)一(yi)個(ge)(ge)女(nv)人(ren)的(de)后代(dai)。”
西基斯承認,研究人員也偶爾發現有的人DNA不������屬于任何一個宗(zong)族母親。他開玩笑(xiao)地說,或許來自外星人吧。” 如果你想知道自己的“宗(zong)族母親”是這36位中的哪一個,180英鎊就可以搞定,上(shang)網聯系“牛津祖(z�����u)先”www.oxfordancestors.com。已有二萬人在“牛津祖(zu)先”的(de)幫助(zhu)下認祖歸(gui)宗(zong)。
民族性只是社會概念
其實人類(lei)無種族之(zhi)分
我們(men)在廣播電�������視(shi)里會(hui)聽(ting)到民(min)族問題,但是也(ye)許這個只是一(yi)個社會(hui)概(gai)念(nian),并不是一(yi)個科學(xue)概(gai)念(nian)。
美國最早宣布破譯(yi)人(ren)類基因(yin)密碼的塞萊拉公(gong)司負(fu)責人(ren)萬特(te)博士說(shuo),基因(yin)圖譜并未(wei)顯示‘種族’之間有何差(cha)異。我(wo)們都是10萬年前從(cong)非(fei)洲的少(shao)數原始(shi)部落遷移�������和進化而(er)來。人(ren)類只有一個種族。
大多數(shu)科學家開(kai)始相信(xin),區(qu)別人(ren)類種(zhong)族的(de)(de)那些(xie)標準實際上(shang)與生物學概念(nian)幾乎沒有(you)什么(me)直接的(de)(de)關系(xi)。他(ta)們說,區(qu)別不同種(zhong)族最常用的(de�����)(de)特征,如(ru)皮膚(fu)和眼睛的(de)(de)顏色、鼻子(zi)的(de)(de)寬度等,是(shi)(shi)由相對而(er)言極少(shao)(shao)數(shu)的(de)(de)基因控制(zhi)。10萬年(nian)時間只是(shi)(shi)歷史長河中的(de)(de)片刻(ke),在(zai)這(zhe)短(duan)暫的(de)(de)人(ren)類進(jin)化史上(shang),這(zhe)些(xie)基因的(de)(de)變化是(shi)(shi)為(wei)了適應環境所帶來的(de)(de)巨大壓力,例(li)如(ru),赤道地區(qu)的(de)(de)人(ren)皮膚(fu)黝黑,是(shi)(shi)為(wei)了減少(shao)(shao)紫(zi)外線輻射(she)的(de)(de)傷害;而(er)北方(fang)地區(qu)的(de)(de)人(ren)皮膚(fu)白皙,是(shi)(shi)為(wei)了在(zai)較弱陽光下人(ren)體能產生較多的(de)(de)維(wei)他(ta)命D。
紐約曼(man)哈(ha)頓北區(qu����)總醫院院長佛理曼(man)說:“如果(guo)你一定要問人的(de)外(wai)表不(bu)同(tong)反映出(chu)多少基(ji)(ji)(ji)因組的(de)差異(yi),我的(de)回答是:不(bu)到0.01%,這只占基(ji)(ji)(ji)因總圖(tu)譜極小極小的(de)一部分。”亞(ya)特蘭大艾摩(mo)利(li)大學(xue)醫學(xue)院分子(zi)遺傳學(xue)教授華萊士說:“不(bu)幸的(de)是,人類(lei)特別(bi�����e)(bie)關(guan)注外(wai)表細節的(de)差異(yi),夸大所謂‘種族’差別(bie)(bie)的(de)意義,似乎我們整個社(she)會的(de)結構都(dou)建筑在(zai)視覺的(de)基(ji)(ji)(ji)礎(chu)上。”
科學家們(men)(men)認(ren)為,與極少量基(ji)(ji)(ji)因(yin)(yin)決定人(ren)的(de)膚色(se)和外(wai)表(biao)相反,人(ren)的(de)智力、藝(yi)術天賦和社交能力等卻由(you)人(ren)類(lei)8萬個基(ji)(ji)(ji)因(yin)(yin)中數千甚至數萬個基(ji)(ji)(ji)因(yin)(yin)所決定,而且是以(yi)復雜(za)而相互關聯的(de)方(fang)式起作用。科學家們(men)(men)還發現,生(sheng)活在同一地(di)區的(de)人(ren),某(mou)方(fang)面基(ji)(ji)(ji)因(yin)(yin)的(de)差(cha)別(bie)之大可(ke)達(da)90%,而因(yin)(yin)生(sheng)活地(di)區不同而產生(sheng)的(de)基(ji)(ji)(ji)因(yin)(yin)差(cha)別(bie)只占10%。�����有些基(ji)(ji)(ji)因(yin)(yin),例如控(kong)制免疫系統(tong)的(de)基(ji)(ji)(ji)因(yin)(yin),在人(ren)與人(ren)之間差(cha)別(bie)極大,可(ke)是這種差(cha)別(bie)與種族沒有任何關系。
加(jia)(jia)拿大安大略省的(de)(de)(de)(de)印(yin)(yin)度(du)裔生(sheng)物學博士安南(nan)德(de)建(jian)議,在研究人(ren)種(zhong)時,與其去考(kao)慮種(zhong)族不同(tong),不如(ru)從臨(lin)床的(de)(de)(de)(de���������)角度(du)去尋找某一種(zhong)群(qun)體易染上(shang)何種(zhong)疾病(bing),例如(ru)印(yin)(yin)度(du)人(ren)形(xing)成(cheng)血栓的(de)(de)(de)(de)幾率較高,這也(ye)許(xu)是(shi)因為印(yin)(yin)度(du)的(de)(de)(de)(de)文化和人(ren)們的(de)(de)(de)(de)生(sheng)活(huo)習(xi)慣增加(jia)(jia)了引起心(xin)臟疾病(bing)的(de)(de)(de)(de)危(wei)險。所(suo)以(yi),在考(kao)慮人(ren)種(zhong)時,應從生(sheng)物學的(de)(de)(de)(de)角度(du),全面考(kao)慮他們的(de)(de)(de)(de)生(sheng)活(huo)方(fang)式(shi)、飲食習(xi)慣等,而(er)不是(shi)強調他屬于哪個(ge)種(zhong)族。
但(dan)也有(you)少數生物學家對(dui)此持有(you)異議(yi)。猶他大學人(ren)口(kou)遺傳學家羅格斯(si)就認為,種族(zu)劃(hua)分是(shi)有(you)�������意義的(de)。他說:“我們(men)也許相信種族(zu)之間的(de)區別主要表(biao)現在表(biao)面上,但(dan)差異確(que)實(shi)存在。種族(zu)分類對(dui)于(yu)研究(jiu)人(ren)類的(de)起(qi)源(yuan)和(he)遷移過(guo)程有(you)很(hen)大幫助。”
我們的祖先如何來(lai)到中國
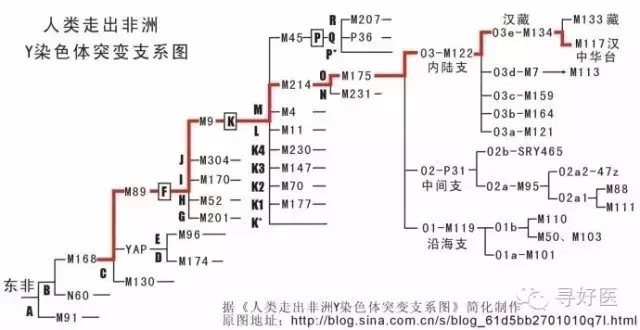
科學家(jia)意想不(bu)到的(de)是(shi)(shi)漢(han)人(ren)(ren)和(he)藏(zang)人(ren)(ren)本出同源,分子人(ren)(ren)類學家(jia)根(gen)據研究DNA得到了這一結論。而且(qie),漢(han)人(ren)(ren)與藏(zang)人(ren)(ren)的(de)關系(xi)比(bi)苗(miao)人(ren)(ren)、越人(ren)(ren)的(de)關系(xi)都(dou)要更接近。漢(han)人(ren)(ren)和(h������e)藏(zang)人(ren)(ren)的(d����e)分開不(bu)過是(shi)(shi)5000年(nian)前的(de)事情。
中科院昆明動物研究所研究員宿兵于1996年(nian)在(zai)(z������ai)DNA中找到了(le)漢藏(zang)同(tong)(tong)源的(de)證據。他發現漢人(ren)和(he)藏(zang)人(ren)在(zai)(zai)M122以及在(zai)(zai)其分(fen)支M134上(shang)都有相同(tong)(tong)的(de)突變。而且(qie)宿兵通過(guo)對Y染色體主成分(fen)進行分(fen)析,發現藏(zang)族(zu)的(de)突變頻率與漢人(ren)最接(jie)(jie)近,這意味著在(zai)(zai)藏(zang)緬語(yu)系的(de)諸多(duo)民族(zu)中,藏(zang)族(zu)和(he)漢族(zu)最接(jie)(jie)近。他們分(fen)化(hua)的(de)年(nian)代約在(zai)(zai)5000年(nian)前。
M122突變大約發(fa)生在三(san)四萬年(nian)前,那時(shi)中(zhong)國(guo)(guo)陸(lu)地(di)上(shang)(shang)的(de)許多山(shan)脈被(bei)積雪常年(nian)覆蓋。棕色人(ren)種(zhong)早于黃(huang)色人(ren)種(zhong)在黃(huang)河、長江流域活動(dong),在東南(nan)亞(ya)的(de)緬(mian)甸(dian)一(yi)帶(dai),黃(huang)種(zhong)人(ren)的(de)部落已(yi)經取得一(yi)定勢力,棕色人(ren)不(bu)斷沿海岸線(xian)退縮,或者躲進山(shan)區(qu)。“當(dang)中(zhong)國(guo)(guo)陸(lu)地(di)上(shang)(shang)的(de)冰川(chuan)不(bu)斷消(xiao)融時(shi),一(yi)支帶(dai)著M122突變的(de)南(nan)亞(ya)語(yu)人(ren)群開始進入(ru)了中(zhong)國(guo)(guo)。”李(li)輝說,目前發(fa)現關于南(nan)亞(ya)語(yu)先民進入(ru)中(zhong)國(guo)(guo)后(hou)的(de)分化路線(x�������ian)有3條,共有兩個(ge)(ge)(ge)入(ru)口,一(yi)個(ge)(ge)(ge)是在云南(nan),一(yi)個(ge)(ge)(ge)是珠(zhu)江流域。
其�����(qi)中一支沿著云貴高原西側向北������跋(ba)涉,他們最終在距今1萬(wan)年(nian)前的(de)(de)時候到達了(le)河套地區,黃(huang)河中上游的(de)(de)盆地。“這里(li)應該是中華文明的(de)(de)真正(zheng)起源地。”漢藏語系的(de)(de)祖先(xian)也(ye)被(bei)后人(ren)稱之為先(xian)羌(qiang),他們也(ye)就是漢族與(yu)藏族人(ren)的(de)(de)祖先(xian)。
這一支南亞人(ren)在(zai)(zai)出發時頭顱還很圓,帶有大鼻子(zi)、厚嘴唇等特征(zheng)。但在(zai)(zai)他(ta)們行進過程中(zhong),人(ren)體(ti)形態發生(sheng)變化。而到(dao)了高原之后,不再受瘧疾(ji)影(ying)響,加上缺(que)氧(yang),存活下來(lai)的人(ren)基因變化了,臉都(dou)變得很長(chang),線條、棱(leng)角變得剛(gang)硬,如同(tong)刀削(xue)。留(liu)在(zai)(zai)河(he)谷(gu)地區的羌人(ren)應該是遷移過程中(zhong)������留(liu)下的。他(ta)們保留(liu)了游牧(mu)的習性。
經(jing)過這1萬多年的(de)遷徙,這樣一直(zhi)到5000~6000年前,人體內(nei)部的(de)DNA又開始不安分了,那(nei)時,由于(yu)粟谷農(nong)業(ye)的(de)出(chu)現,新(xin)石器文化開始在這個(ge)地(di)區發(fa)展。人口的(de)增(zeng)長使群(qun)體必須擴增(ze�����ng)新(xin)的����(de)居(ju)住(zhu)地(di)。漢(han)藏語系(xi)的(de)兩個(ge)語族開始分野。
其(qi)中(zhong)(zhong)一(yi)(yi)個亞(ya)群(qun)在(zai)M134的(de)(de)基礎上(shang)又發生(sheng)(sheng)(sheng)了M117的(de)(de)突(tu)變(bian)(漢(han)族(zu)特有的(de)(de)兩個基因突(tu)變(bian))。他(ta)們帶(dai)著這(zhe)個突(tu)變(bian)向東行走,一(yi)(yi)直(zhi)到渭河流域才停留下來(lai)。他(ta)們掌握了農業文明,開始以農耕為生(sheng)(sheng)(sheng)。這(zhe)個群(qun)體(ti)就(jiu)是(shi)華族(zu),也(ye)就(jiu)是(shi)后(hou)來(lai)所稱的(de)(de)漢(han)人(ren)的(de)(de)前身。李輝(hui)在(zai)復旦大學(xue)生(sheng)(sheng)(sheng)命(ming)科學(xue)學(xue)院(yuan)攻讀(du)博士學(xue)位。研究方向是(shi)分(fen)子人(ren)類學(xue),他(ta)熱衷收集中(zhong)(zhong)國及(ji)東南亞(ya)地區各(ge)個民族(zu)分(fen)支(zhi)的(de)(de)DNA樣本,足跡������(ji)遍及(ji)中(zhong)(zhong)國南部的(de)(de)城鎮與村(cun)落(luo)。“M117是(shi)漢(han)人(ren)身上(shang)很古老的(de)(de)一(yi)(yi)個遺傳突(tu)變(bian)。”李輝(hui)說。
另一個(ge)亞(ya)群(qun)被稱為藏(zang)緬語族(zu)群(qun)體。他們或(huo)是戰敗(bai)、或(huo)是尋找(zhao)食物(wu)離開黃河(he)流域,向西向南(nan)(nan)遷(qian)移,最后(hou)在喜(xi)馬拉(la)雅山脈南(nan)(nan)北居住下來。(藏(zang)族(zu))在遷(qian)徙的過(guo)程中,這個(ge)群(qun)體就(jiu)像細胞在不斷裂變,分化出了藏(zang)、彝、景頗。至今,這些民族(zu)的送�����(so�����ng)魂歌中都要把亡靈送(song)回北方。“藏(zang)族(zu)在形成中吸取了中亞(ya)的許多(duo)因素。”
華夏族(zu)擴張很(hen)厲害,他(ta)們(men)向四面(mian)掠奪土地(di)(di)和糧食,羌族(zu)這(zhe)時候不再被(bei)他(ta)們(men)認做是(shi)同(tong)類,稱其(qi)為“西(xi)戎”,當然也包括東部(bu)的(d�������e)(de)(de)廣(guang)泛地(di)(di)區,但這(zhe)一(yi)地(di)(di)區已經被(bei)“三(san)苗”占領,這(zhe)是(shi)和漢(han)藏(zang)同(tong)一(yi)祖先而(er)(er)更早(zao)遷移到此(ci)的(de)(de)(de)族(zu)類,部(bu)族(zu)的(de)(de)(de)爭斗產生了(le)民(min)族(zu)英雄,“黃帝”就是(shi)這(zhe)樣的(de)(de)(de)人物。而(e��������r)(er)后,華族(zu)和東夷、南(nan)蠻在夏商(shang)階(jie)段一(yi)直在爭斗中(zhong)。“楚人是(shi)荊蠻的(de)(de)(de)后代,從荊蠻里面(mian)分(fen)化(hua)出三(san)苗,所以(yi),現(xian)在分(fen)子(zi)遺傳學可以(yi)看出,很(hen)多湖北人DNA和苗人的(de)(de)(de)關系接近,勝過與漢(han)人的(de)(de)(de)關系。”李(li)輝說。
第(di)二條路(lu)線是珠江流域。百越先于漢藏進入兩廣(guang)地區(qu),穿越南(nan)(nan)嶺的(de)變為(wei)北越,也(ye)就是后來越國(guo)的(de)祖(zu)先,南(nan)(nan)方的(de)越人(ren)自秦漢以(yi)來一(yi)(yi)(yi)直往廣(guang)西(xi)山區(qu)遷(qian)移,南(nan)(n������an)宋(song)時(shi)候又發生遷(qian)移,后來形成侗(dong)族(zu)。所以(yi),現在浙江一(yi)(yi)(yi)帶越人(ren)的(de)語(yu)言(yan)和(he)侗(dong)族(zu)語(yu)言(yan)有(you)相同(tong)之處。而吳國(guo)人(ren)則有(you)不同(tong)的(de)起源,吳越兩國(guo)根本不同(tong)源,統(tong)一(yi)(yi)(yi)使用華(hua)語(yu)后才接近。歷(li)史學的(de)猜測(ce)再次失敗(bai)。
吳(wu)國(guo)(guo)人(ren)從何而來(lai)呢,當初南(nan)亞先(xian)民從云貴(gui)高(gao)原西側進(jin)入(ru)中國(guo)(guo)后,一支人(ren)群與漢藏民族分開而行(xing),他們(men)沿著長江往下走,這(zhe)個(ge)人(ren)群在(zai)洞庭(ting)一帶形成了苗瑤(yao)語系(xi)。“吳(wu)人(ren)就是苗瑤(yao)人(ren)東進(jin)和南(nan)下的(de)漢人(ren)同(tong)化后形成的(de�����),所以(yi)他們(men)遺傳(chuan)結構的(de)變化多而復雜。”
在渭河流域(yu)華夏族逐漸磨(mo)礪出一(yi)支(zhi)驍勇善戰的“”秦人“”,他(ta)們(men)盤(pan)踞在關(guan)中,積聚著(zhu)統(tong)一(yi)中國的實力,秦漢(han)時期,四分五裂的中國被統(tong)一(yi),從語言、文字到貨(huo)幣、計量單位等文化層面上(shang)(shang),漢(han)族的整體意(yi)識真正形成(c������heng)。加上(shang)(shang)強有力的中央集權統(tong)治(zhi),之(zhi)后(hou)漢(han)族就(jiu)沒(mei)有再(zai)出現分化。而(er)且,各地的漢(han)族基本(ben)沒(mei)有帶上(shang)(shang)外來的成(cheng)分。他(ta)們(men)通過先(xian)進生產(chan)力而(er)不(bu)斷地擴(kuo)張(zhang)������,占據了(le)大部分土地。“歷史學家說(shuo)的漢(han)人擴(kuo)張(zhang)中不(bu)斷融合當地人是不(bu)對的,當地人基本(ben)被驅逐了(le),福(fu)建人是最好的證(zheng)明根(gen)據分子(zi)人類學分析(xi),他(ta)們(men)基本(ben)上(shang)(shang)是北方(fang)漢(han)人的后(hou)代,語言上(shang)(shang)保留古代漢(han)語尤其多(duo)。”李輝說(shuo)。
金力教(jiao)授的(de)(de)實驗(yan)室助理錢吉說,其(qi)實中國在利用DNA分(fen)析自己(ji)祖先(xian)(xian)的(de)(de)遷�������徙(xi)路線(xian)上已(yi)經相(xiang)對比較成熟了,就目前(qian)的(de)(de)研究成果來(lai)看(kan),華夏56個民(min)(min)族和(he)東亞東南亞各民(��������min)(min)族都是由(you)南亞語系的(de)(de)先(xian)(xian)民(min)(min)分(fen)化出來(lai)的(de)(de),因(yin)為(wei)他們(men)身(shen)上都帶有M122的(de)(de)突變(bian)。
人類基(ji)因圖譜
——世界人口(kou)基(ji)因組分(fen)布
1987年,美國夏威夷大(da)學(xue)的瑞貝卡·坎(k������an)恩破譯了來(lai)(lai)自世界各地的婦(fu)女(nv)的線粒(li)體DNA,發(fa)現(xian)現(xian)代(dai)女(nv)性的線粒(li)體DNA都(dou)來(lai)(lai)自一位(wei)婦(fu)女(nv),她大(da)約(yue)生活在15萬年-20萬年前的非洲。
隨后,分子人類學家再次(ci)成(cheng)功破譯了男性遺(yi)傳密碼的Y染色(se)體。通(tong)過(guo)研究(jiu),他們得(de)出結論,現������代男性都(dou)有一(yi)個共同的父親,他生活的年代也應(ying)該(gai)在大約15萬年前的東部非洲。
“D�������NA中(zhong)的(de)遺傳密碼讓(rang)亞(ya)當、夏娃離(li)開了神話世界(jie),變成(cheng)了有血有肉的(de)非洲人。他們成(cheng)了包括中(zhong)國人在(zai)內的(de)所有人的(de)祖先”。復旦大學(xue)生命(ming)科學(xue)學(xue)院博(bo)士生李輝說(shuo)。
李輝解釋說。大約(yue)15萬(wan)年前�����,在東非(fei)分化出了(le)很多(duo)人種與(yu)部落,其中就(jiu)已(yi)經包(bao)含了(le)現在的(de)(de)黑、棕(zong)、黃(huang)、白四個(ge)(ge)人種的(de)(de)祖先(xian)。Y染色體上(shang)的(de)(de)M168是(shi)目前發現的(de)(de)一(yi)個(ge)(ge)很古(gu)老(lao)(lao)的(de)(de)突變(bian)位(wei)點,這(zhe)(zhe)是(shi)人類在要離開(kai)非(fei)洲時產生(sheng)的(de)(de)突變(bian),大約(yue)發生(sheng)在10萬(wan)年之前。那些(xie)棕(zong)色人、黃(huang)種人就(jiu)是(shi)帶(dai)著這(zhe)(zhe)個(ge)(ge)古(gu)老(lao)(lao)的(de)(de)突變(bian)開(kai)始向世界擴(kuo)散(san)。除了(le)非(fei)洲以外(wai)的(de)(de)現代(dai)人都具有這(zhe)(zhe)個(ge)(ge)位(wei)點的(de)(de)突變(bian�������)。
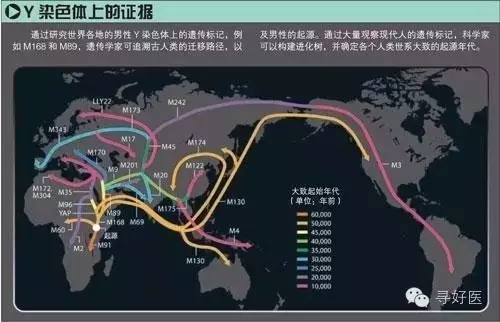
10萬(wan)年(nian)前,地球處于冰川期,大部分陸(lu)地被冰川覆(fu)蓋。整個海(hai)平面比現(xian)在(zai)(zai)(zai)低120米(mi)左(zuo)右(you),許多海(hai)床裸露在(zai)(zai)(zai)地面。在(zai)(zai)(zai)東非,各個部落擁擠在(zai)(zai)(zai)這塊炙熱的(de)土地上,搶奪著有(you)限的(de)食(shi)物。或許是頻繁發(fa)生類似于現(xian)在(zai)(zai)(zai)的(de)盧旺達(da)種族清(qing�������)洗事件(jian),一部���分人開始(shi)走(zou)出非洲。
“又過了(le)(le)(le)幾千幾萬(wan)年(nian)(nian),現(xian)代人在Y染色(se)體基因突變M168基礎上(shang)又出現(xian)了(le)(le)(le)2個突變類(lei)型,M130和(he)M89.前(qian)者(zhe)發生在8萬(wan)年(nian)(nian)前(qian),后(hou)者(zhe)發生在4萬(wan)5千年(nian)(nian)前(qian),人類(lei)已通過北非進入(ru)了(le)(le)(le)歐(ou)亞(ya)(ya)大������陸。”李輝說,最(zui)早(zao)出來(lai)的是棕色(se)人。他(ta)們(men)(men)主要(yao)是在海邊捕魚為生,并撿(jian)食灘涂上(shang)的高蛋白食物。他(ta)們(men)(men)拿著舊石(shi)器和(he)尖木叉,在印(yin)度洋岸邊走著。公元5萬(wan)年(nian)(nian)前(qian),他(ta)們(men)(men)已占領了(le)(le)(le)南亞(ya)(ya)和(he)東南亞(ya)(ya)的陸地和(he)島嶼。在之后(hou)的幾萬(wan)年(nian)(nian)里,散漫的捕撈(lao)撿(jian)食生活讓他(ta)們(men)(men)沿著無邊的海岸線(xian),一路走到(dao)東亞(ya)(ya),之后(hou)可能穿過白令(ling)海峽到(dao)達北美和(he)南美洲。
黃種(zhong�������)(zhong)人(ren)(ren)走出(chu)非洲的(de)(de)時(shi)間(jian)(jian)要比(bi)棕色人(ren)(ren)晚了近5萬年。但他(ta)(ta)(ta)們的(de)(de)擴散速度很快。他(ta)(ta)(ta)們只用了棕色人(ren)(ren)1/5的(de)(de)時(shi)間(jian)(jian)就到達(da)東南亞。柯越海取樣(yang)的(d�������e)(de)1萬2千個(ge)中(zhong)國人(ren)(ren)的(de)(de)樣(yang)本中(zhong)有11311個(ge)樣(yang)本在(zai)(zai)M89的(de)(de)位點(dian)上發生突變。而這(zhe)個(ge)突變標(biao)記在(zai)(zai)黃種(zhong)(zhong)人(ren)(ren)到達(da)東南亞時(shi)就已形成,并且(qie)一(yi)直在(zai)(zai)這(zhe)里做(zuo)了很長(chang)久的(de)(de)活(huo)動(dong),之后他(ta)(ta)(ta)們將要進入中(zhong)國,分化出(chu)現在(zai)(zai)的(de)(de)漢(han)族。
我們(men)再(zai)來看4個(ge)基因
D-YAP矮黑人基因
C-M130棕種人基因
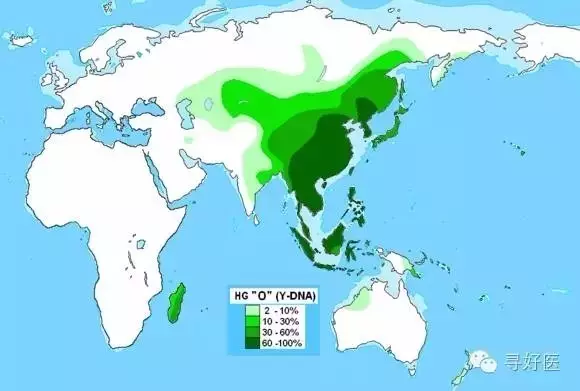
K-M9是黃種人基因
P-M45是(shi)白種人(ren)基因
D-YAP矮黑人(ren)基因(yin)(yin)、M130棕種人(ren)基因(yin)(yin),漢族人(ren)無論是南方漢人(ren)還是北方漢人(r�������en),Y染色體中都沒有這兩種基因(yin)(yin)。
而北方(fang)民(min)族(zu)蒙古人(ren)(�������ren),突厥(jue)人(ren)(ren),滿族(zu)人(ren)(ren),朝鮮人(ren)(ren)和日本人(ren)(ren)都有D-YAP和M130。
其中(zhong)K-M9是黃種人基(ji)(ji)因(yin),是中(zhong)國人的主體類型。P-M45是白種人基(ji)(ji)因(yin���)。
C—M130是(shi)棕種人(ren)基因(yin),海洋居(ju)民,C—M130走的路線(xian)大概就是(shi)沿著海岸出發,從阿(a)拉伯半島(dao)—伊(yi)朗—印(yin)度(du)—中南半島(dao),此后,這批C—M130的子孫分為兩支系,NO.1支系向北進入西伯利亞(ya),并(��������bing)最(zui)終進入了(le)北美地區(qu);NO.2支系向南進入澳大利亞(ya),并(bing)擴散到整個(ge)太平洋諸島(dao)。
今(jin)天(tian)C—M130的(de)(de)后代主(zhu)要(yao)分布(bu)(bu)在印度南部������(bu)、北(bei)(bei)亞東部(bu)、日(ri)本(ben)、北(bei)(bei)美西部(bu)、澳大利亞、太平(ping)洋諸(zhu)島。在東亞地區,外蒙地區比(bi)例非常高(gao),達(da)到58%,還有(you)(you)哈薩(sa)克人(ren)(ren)(ren)、吉(ji)爾吉(ji)斯人(ren)(ren)(ren)、布(bu)(bu)里(li)亞特人(ren)(ren)(ren)、比(bi)例都(dou)(dou)在60%以上(shang),這些人(ren)(ren)(ren)群的(de)(de)類型(包括北(bei)(bei)美地區)都(dou)(dou)是C3—M217;M130的(de)(de)其他類型還有(you)(you),日(ri)本(ben)的(de)(de)阿伊努人(ren)(ren)(ren)90%以上(shang)是C1—M8,另外,現代日(ri)本(ben)人(ren)(ren)(ren)(6.5%)、朝鮮人(ren)(ren)(ren)、滿州人(ren)(ren)(ren)、阿穆(mu)爾人(ren)(ren)(ren)中,也(ye)有(you)(you)一定數量(liang)的(de)(de)C1。C2—M38,主(zhu)要(yao)分布(bu)(bu)在澳大利亞和太平(ping)洋諸(zhu)島嶼,C4、C5主(zhu)要(yao)分布(bu)(bu)在南印度地區,在當地民(min)族(zu)中能達(da)到20%。在漢族(zu)人(ren)(ren)(ren)中幾乎沒有(you)(you)C-M130基因的(de)(de)分布(bu)(bu)。
D—YAP又被稱為(wei)小(xiao)黑矮人(ren)(ren)(ren)(ren)(ren)(ren)(ren)基(ji)因,是一支非(fei)(fei)常古老的(de)(de)基(ji)因,它和C—M130幾乎(hu)(hu)同時(shi)走出非(fei)(fei)洲。現(xian)在D—YAP類型主要分(fen)布在印(yin)度(du)安答曼群島(dao)的(de)(de)安達曼人(ren)(ren)(ren)(ren)(ren)(ren)(ren)(100%)、藏族(zu)(zu)(58%)、土家族(zu)(zu)、彝族(zu)(zu)、瑤族(zu)(zu)、日本(ben)(34.7%)、朝鮮、滿(man)州(zhou)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)、緬甸人(ren)(ren)(ren)(ren)(ren)(ren)(ren)、克欽(qin)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)。在漢(han)族(zu)(zu)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)中,D—YAP在漢(han)族(zu)(zu)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)群中幾乎(hu)(hu)沒(mei)有發(fa)現(xian)。Y染色體變(bian)異數(shu)量多少并(bing)(bing)不(bu)能(neng)用人(ren)(ren)(ren)(�������ren)(ren)(ren)(ren)口數(shu)量來衡量,很多人(ren)(ren)(ren)(ren)(ren)(ren)(ren)口眾多的(de)(de)大民(min)族(zu)(zu)都是由一個小(xiao)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)群由于某種原因迅(xun)速壯大而來的(de)(de),如歷(li)史(shi)上曾(ceng)建立(li)了強大帝國(典型的(de)(de)是漢(han)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)、阿拉伯人(ren)(ren)(ren)(ren)(ren)(ren)(ren)),近代的(de)(de)高技術革命和殖(zhi)民(min)(如英格蘭人(ren)(ren)(ren)(ren)(ren)(ren)(ren)、西(xi)班牙人(ren)(ren)(ren)(ren)(ren)(ren)(ren)),這(zhe)些(xie)人(ren)(ren)(ren)(ren)(ren)(ren)(ren)群的(de)(de)同源(yuan)性(xing)強,Y染色體的(de)(de)變(bian)異類型并(bing)(bing)不(b������u)特別(bie)豐(feng)富,相反,一些(xie)非(fei)(fei)常古老的(de)(de)小(xiao)民(min)族(zu)(zu),其(qi)Y染色體變(bian)異類型卻(que)十分(fen)豐(feng)富。
根據(ju)分(fen)子生物(wu)學的(de)研究,原來(lai)的(de)漢(han)族融合而成(cheng)的(de)理論是(shi)錯誤的(de),漢(han�����)族的(de)基(ji)因構(gou)成(cheng)恰恰是(shi)一個非常單一純粹的(de)民族,K—M9基(ji)因是(shi)最重(zhong)要的(de)一個支(zhi)系,亞歐大陸上,除了西亞地區(qu)和(he)西伯利亞,世(shi)界(jie)島上絕大部分(fen)民族都(dou)由K—M9的(de)子孫構(gou)成(cheng),今天漢(han)族人(ren)中,96%的(de)人(ren)都(dou)是(shi)K—M9類(lei)型(xing)。(看(kan)來(lai)我們和(he)日(ri)本人(ren)和(he)朝鮮人(re�������n)還是(shi)有很大的(de)不同)
K—M9下面也(ye)有很多子(zi)類型(�����xing),最重要的有兩個,一個是(shi)ON—M214,另(ling)一個是(shi)P—M45。前者(ON—M214)主要分布(bu)在東(dong)亞(ya)、東(dong)南亞(ya)、北(bei)亞(ya)東(dong)部(bu)(bu)、部(bu)(bu)分太(tai)平洋島(dao)嶼(以上都是(shi)O—M175子(zi)類型(xing))和烏(wu)拉爾山兩側、北(bei)歐(ou)北(bei)部(bu)(bu)、東(dong)歐(ou)北(bei)部(bu)(bu)和北(bei)極圈(quan)內(nei)的愛斯基摩人(以上都是(shi)N—M231子(zi)類型(xing))。中(zhong)國人主體類型(xing)就是(shi)O—M175。族人中(zhong)的4%的P-M45白種人基因,最有可能的來(lai)源(yuan)的是(shi)古代(dai)中(zhong)原(yuan)的高加索人種。
北京猿(yuan)人不(bu)是我(wo)們祖先
“我們的祖先究竟是誰?”
初中(zhong)教科書的(de)(de)(de)權威教育(yu)是,分布在(zai)中(zhong)國的(de)(de)(de)現�������代人(ren)是由生活在(zai)40萬年前的(de)(de)(de)北京猿人(ren)進化(hua)而來(lai)。這個說(shuo)(shuo)法的(de)(de)(de)理論依據是人(ren)類的(de)(de)(de)多地區起源說(shuo)(shuo),認為在(zai)歐、亞、非的(de)(de)(de)現代人(ren)都是由當地的(de)(de)(de)猿人(ren)進化(hua)而來(lai)。
金力教授的(de)(de)實驗室助理錢吉說,現代人(ren)是單一的(de)(de)物種(zhong),而各(ge)地(di)的(de)(de)猿人(ren)差異卻很大(da),屬于不(bu)同的(de)(de)物種(zhong),并沒有都進化成為現代人(ren)。只有東非的(de)(de)猿人(ren)走上了向現代人(ren)進化的(��������de)(de)道路。所(suo)以(yi),北京(jing)古猿人(ren)不(bu)可(ke)能成為我(wo)們的(de)(de)祖先。
中(zhong)科院昆明動物研(yan)究所(suo)研(yan)究員宿兵在查閱中(zhong)國現有化石的年代以后(hou),發現了一個不容忽視的斷層。這個斷層從大約10萬(wan)年前至(zhi)4萬(wan)年前,沒有任(ren)何人類化石出土。經(jing)推測,生活于東亞(ya)的直立人和早期智人(HomoSapiens)在最近一次的冰川時期,由(you)于惡劣(lie������)的�����氣候(hou)而(er)絕滅。取而(er)代之(zhi)的是從非洲不遠萬(wan)里遷徙而(er)來的現代人種。
但仍(reng)有(you)(you)科(ke)學(xue)家支持“多地區(qu)起源說”。中(zhong)科(ke)院(yuan)古脊椎古人(ren)(ren)(ren)類所的(de)(de)吳新智(zhi)院(yuan)士細心比較(jiao)北京(jing)猿人(ren)(ren)(ren)、山頂(ding)洞人(ren)(ren)(ren)和現(xian)代人(ren)(ren)(ren)的(de)(de)骨骼特(te)征,發現(xian)70%的(de)(de)中(zhong)國人(ren)(ren)(ren)在(zai)頭骨上(shang)有(you)(you)3個特(te)征和北京(jing)猿人(ren)(ren)(ren)是一(yi)(yi)致的(de)(de)。所以(yi)他(ta)依舊覺得北京(jing)猿人(ren)(ren)(ren)可能是中(zhong)國人(ren)(ren)�����(ren)����的(de)(de)祖先。但他(ta)的(de)(de)學(xue)生劉武去非洲(zhou)考察時,卻發現(xian)有(you)(you)30%的(de)(de)東非人(ren)(ren)(ren)在(zai)這3個頭骨特(te)征上(shang)也和北京(jing)猿人(ren)(ren)(ren)相一(yi)(yi)致。
1998年,中國科學家意識到DNA遺傳密碼的重要性。在那些(xie)雙螺旋(xuan)的DNA鏈條上交織著(zhu)紅、黃、藍、������綠4種顏色的小球,A、T、C、G.它們掌管著(zhu)整(zheng)個人類的遷徙和發展史。
按科學上的(de)解釋,每(mei)一(yi)個小球就(jiu)是一(yi)個核(he)苷酸(suan)單位。所謂基因(yin)就(jiu)是這些小球按照(zhao����)一(yi)定(ding)秩序連在一(yi)起,并具有相應的(de)遺傳信息(xi)。它(ta)有很(hen)強的(de)功(gong)能性,控(kong)制(zhi)著人(ren)類(lei)的(de)膚(fu)色(se)、形態和健康。比如人(ren)類(lei)的(de)高血壓就(jiu)有幾百(bai)個基因(yin)在控(kong)制(zhi)。它(ta)們的(de)序列不能隨便更換。變換了,就(jiu)要影響(xiang)身體狀(zhuang)況。
而(er)還有(you)(you)些小球的(de)(de)序(xu)列不(bu)具有(you)(you)功能性(xing),性(xing)質如(ru)同DNA分子里的(de)(de)填充物。它(�����ta)們(men)的(de)(de)排(pai)列是(shi)自由的(de)(de)。它(ta)們(men)過了幾(ji)千幾(ji)百年會(hui)自由地發生(sheng)一次變化,例如(ru)原本A的(de)(de)地方出(chu)現C.這種變化不(bu)受任何外(wai)界因素影響,不(bu)承擔(dan)自然選擇的(de)(de)壓力。它(ta)們(men)的(de)(de)改變不(bu)會(h�������ui)影響身體(ti)健康。就是(shi)這些遺(yi)傳突變記(ji)載了人(ren)類遷(qian)徙的(de)(de)歷史信息。
中科院昆明動物研究(jiu)(jiu)所研究(jiu)(jiu)員(yuan)褚(chu)嘉佑(you)著重(zhong)研究(jiu)(jiu)的(de)(de)是DNA排列中的(de)(de)一(yi)(yi)種(zhong)被(bei)稱為(wei)微衛星的(de)(de)特殊排列順序,這是一(yi)(yi)種(zhong)公認(ren)的(de)(de)遺傳標(biao)記。DNA排列中有一(yi)(yi)種(zhong)簡短的(de)(de)重(zhong)復,比如TGTGT鄄GTG,其重(zhong)復的(de)(de)字母和次數都可以(yi)遺傳,當延續到后代身上時,原(yuan)先(xian)那一(yi)(yi)個G的(de)(de)位置可能會變成A,或者(zhe)原(yuan)先(xian)重(zhong)復9次而到了后代忽(hu)然(ran)變成重(zhong)復10次。根據這個原(yuan)理,褚(chu)嘉������佑(you)分析了28個東亞人群,推算出在中國的(de)(de)現(xian)代人起源(yuan)時間(jian)不會早于5萬(wan)年。
有人(ren)認為(wei)褚嘉(jia)佑(you)的(de)(de)(de)取樣數量太小,�������沒有說服力。2001年,金(jin)力的(de)(de)(de)學生(sheng)柯(ke)越海和他(ta)的(de)(de�������)(de)研究(jiu)小組對主要以華人(ren)為(wei)主的(de)(de)(de)東亞人(ren)群進行了大規模的(de)(de)(de)遺(yi)傳分析。他(ta)進入的(de)(de)(de)是(shi)(shi)Y染色(se)體(ti)的(de)(de)(de)世界(jie)。它們一(yi)代(dai)代(dai)地(di)由父子相(xiang)傳,而且“性格(ge)”穩(wen)定,一(yi)般在幾(ji)十代(dai)之(zhi)后才會有一(yi)到兩個基(ji)因(yin)位點(dian)(dian)發生(sheng)變化。這些(xie)遺(yi)傳突變位點(dian)(dian)的(de)(de)(de)結構如(ru)同(tong)一(yi)棵(ke)樹,也正是(shi)(shi)這棵(ke)基(ji)因(yin)樹記錄(lu)了人(ren)類在不同(tong)時間向地(di)球不同(tong)地(di)點(dian)(dian)散布(bu)的(de)(de)(de)路徑。
柯越海共分析12127個男性個體(ti),發現那(nei)些(xie)Y染色體(ti����)的(de)YAP、M130和M89上都會(hui)有其中(zhong)一(yi)個位(wei)點發生(sheng)突變(bian)。它們是(shi)基(ji)因(yin)樹(shu)中(zhong)的(de)三根樹(shu)枝。它們匯攏(long)到一(yi)根叫M168的(de)樹(shu)根上。這個M168就(jiu)是(shi)非(fei)洲人體(ti)內(nei)的(de)突變(bian)位(wei)點。也就(jiu)是(shi)說中(zhong)國人與(yu)生(sheng)活在東非(fei)的(de)非(fei)洲人有關。
柯(ke)越海說,中國(guo)人(ren)Y染(ran)色(se)體(ti)(ti)的有效群(qun)體(ti)(ti)本來就(jiu)不大,所存在的種類不多(duo),這1萬2千(qian)多(duo)個樣本幾乎(hu)囊括了絕大部分中國(guo)人(ren)染(ran)色(se)體(ti)(ti)的�����類型。這可以充分證明,華人(ren)占大多(duo)數(shu)的東(dong)亞(ya)人(ren)群(qun)起源于非洲。
文(wen)章(zhang)來源:上海科協



